植物在自然生活环境中面临着诸多如极端温度,干旱和高盐度等非生物胁迫影响。与生物胁迫不同, 非生物胁迫会引起植物体内多个基因或途径共同发挥作用。虽然多个参与植物对非生物胁迫的基因和信号网络已经被鉴定,但通过改变单一效应基因的表达来提高植物抵抗非生物胁迫能力的研究进展往往不尽人意(Cabello et al., 2014)。天冬酰胺链接的糖基化(Asn-linked glycosylation, N-糖基化)是真核生物中一种重要的蛋白共翻译和翻译后修饰方式。在内质网中,三分支的N-糖基结构在寡糖基转移酶的作用下被转移到新生肽链Asn-X-Ser/Thr motif中的天冬酰胺残基上。随着蛋白折叠由内质网分泌到高尔基体,N-糖基上的甘露糖会在甘露糖苷酶和其他糖原转移酶的作用下被剪切和修饰形成复杂糖基。研究发现绝大部分分泌蛋白或者膜蛋白都在分泌途径中进行N-糖基化。近年来,越来越多的实验证据表明N-糖基在高尔基体内的复杂化对于植物抵抗盐胁迫具有重要的功能(Kang et al., 2008;Strasser et al., 2016),而通过对功能基因的克隆发现在盐土植物中也存在着相似的糖基化过程(Jiang et al., 2015),但N-糖基化参与植物抵抗盐胁迫的分子和生化机制还了解甚少。
2月1日,伟德BETVlCTOR1946洪治研究团队在国际知名学术期刊Molecular Plant上在线发表题为“Trimming of N-glycan by Golgi a1,2 Mannosidases MNS1 and MNS2 is Crucial for Maintaining RSW2 Protein Abundance Against Salt Stress in Arabidopsis”的研究论文,论文链接为http://www.cell.com/molecular-plant/fulltext/S1674-2052(18)30026-1。该研究利用遗传杂交的方法获得了拟南芥高尔基体I型甘露糖苷酶(MNS1和MNS2)功能缺失双突变体,并发现在盐胁迫条件下双突变体mns1 mns2会产生严重的根系生长抑制。有趣的是,缺少C支链的N-糖基生物合成缺陷突变体却表现出耐盐性并且可以部分回复mns1 mns2双重突变体的盐敏感表型;而野生型植株在甘露糖苷酶抑制剂Kifunensine处理下也显示出相似的盐敏感表型,因此推断N-糖基C分支上的a1,2-甘露糖残基的存在是mns1 mns2突变体盐敏感性的“罪魁祸首”,N-糖基C支链的a1,2-甘露糖基的剪切对拟南芥抵御高盐逆境至关重要。该研究还通过生化检测发现盐敏感突变体的根尖纤维素合成受损严重,并进一步研究验证了纤维素合成关键酶--b1,4-葡聚糖酶RSW2的蛋白丰度受C分支上的a1,2-甘露糖残基调控(见附图1)。据此推测,高尔基体中N-糖基的剪切和复杂化的生物学功能是产生一系列寡糖信号,用于细胞内的分子识别和细胞表面蛋白功能状态的调节,这种调节为帮助植物应对胁迫环境至关重要,而这些信号叠加最终形成了复杂型N-糖基。
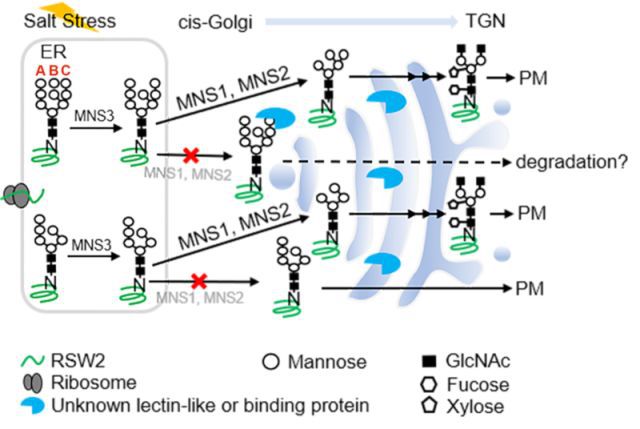
(图1)MNS1/MNS2介导的N-糖基剪切作用调节盐胁迫下RSW2/rsw2-1的稳定性和/或运输机制示意图。
C-分支a1,2-甘露糖残基的切除确保了RSW2/rsw2-1向反式高尔基体网络(TGN)和质膜(PM)的成功运输。而MNS1和MNS2功能的缺失使RSW2/rsw2-1蛋白N-糖基的 C-末端携带a1,2-甘露糖残基,其可能被高尔基体中的某些糖结合蛋白识别并被递送至液泡或其他亚细胞结构或在未知的降解机制中进行降解。
该研究表明蛋白质的翻译后修饰-N-糖基化在调节糖蛋白丰度并维持植物细胞耐盐性中扮演重要角色,提示我们可能通过改变细胞内整体的蛋白质糖基化水平或者N-糖基结构改良植物的耐盐能力,为了解植物对高盐等非生物胁迫的响应机制提供了新的线索,也为采用新方法培育具有优良耐盐性的现代农作物及其分子设计提供了理论基础。
该论文的通讯作者为洪治教授和李建明教授,第一作者为博士研究生刘传发和牛冠婷。相关工作得到了国家自然科学基金优秀青年基金和国家重点基础研究发展计划资助。